Bryopsidaceae
zum Glossar mit Abbildungsverweisen über:

Bryopsidaceae, Moosalgen
1 Wenig, was zusätzlich wichtig erscheint
.
Fortpflanzungszellen[1] entsteh‘n in spezialisierten Zellen
Des am Leben bleibenden Thallus‘[2];
Separate Zonen der Moosalge werden dafür genommen
.
Viele, doch nicht alle Arten bilden
.
Zellwände bauen sie hauptsächlich
.
Fußnoten
[1] Gameten oder Zoosporen
[2] Thallus: Körper von Pflanzen (Pilzen), die nicht in Spross, Blatt und Wurzel gegliedert sind
[3] Sporangium: Behälter, in dem Sporen asexuell oder sexuell gebildet werden
[4] Gametangien: Behälter für Gameten; oder für Zellkerne, die, ohne Gameten zu bilden, dazu bestimmt sind, früher oder später über Karyogamie der sexuellen Fortpflanzung zu dienen
[5] Stephanosporen: Zoosporen mit subapikalem Kranz aus Geißeln
[6] Lebenskreislauf, Lebenszyklus, Entwicklungszyklus, Entwicklungskreislauf: Ein Kreislauf (eigentlich ist es eine Schraube, weil immer Neues entsteht, das zwar dem Anfänglichen gleicht, doch zeitlich später kommt), in dem sexuelle Fortpflanzung erfolgt. Zusätzlich kann noch in regelmäßigem Wechsel eine Phase der asexuellen Vermehrung eingeschlossen sein.
[7] Mannan: Polymer aus β-1,4 glycosidisch verknüpften Mannose-Einfachzucker
[8] Cellulose: Unverzweigte Ketten aus Glucose in β-1,4-Verknüpfung; wobei der 6C-Zucker Glucose in Ring-Form geschrieben, das C1 der Aldehydgruppe ist [CH2OHCHOHCHOHCHOHCHOHCHO], davon aus gerechnet ist der vierte Kohlenstoff das C4 ist. In Ringform geschrieben weist die OH-Gruppe des C1 nach oben, wie auch die frei gebliebene CH2OH-Gruppe. Die OH-Gruppen wechseln von 1 bis 4 die Stellung: C1 nach oben, C2 nach unten, C3 noch oben, C4 nach unten, an C5 hängt die nach oben stehende CH2OH-Gruppe.
[9] Xylan (Bryopsidales): Polymer aus β-1,3 glycosidisch verknüpften Xylose-Einheiten
Eingestellt am 21. Februar 2026
.
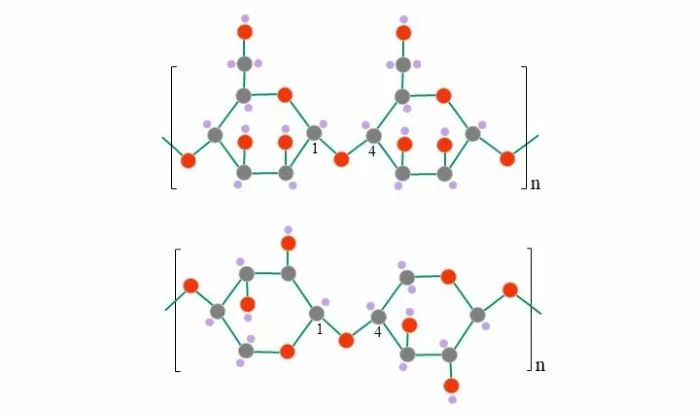
Mannan und Xylan (ppt-generiert; Reinhard Agerer)
Oben: Mannan; β-1,4 verknüpfte Mannose
Unten: Xylan; β-1,4 verknüpfte Xylose (reines Xylan, das nur aus Xylose verknüpft ist; Xylane im weiteren Sinne können verschiedene Seitenfunktionen tragen)
Grau: Kohlenstoff; rot: Sauerstoff; violett: Wasserstoff; dünne grüne Linie: Einfachbindung
Eingestellt am 21. Februar 2026
.

Bryopsidaceae, Moosalgen i.e.s.
2 Einfach organisiert
.
Wie Büschel steh’n sie in Meeren nach oben,
Oder als Federn am spärlich nur Äste tragenden, mageren Stamm.
Strauch wie Baum entspringen verzweigten Stolonen[1],
Die kriechend mit Rhizoiden[2] sich halten am Grund.
.
Verzweigen sich mehrfach, bekleiden aber sie nicht;
Halten allein sich mit dicken Wänden der Schläuche
Ohne zu knicken im wellenbewegten Meer.
.
Bilden Gametangien[5] an Enden von Zweigen begrenzter Areale,
Schotten mit Septum[6] als separate Behälter sie ab.
Erscheinen dunkelgrün, werden große, weiblich gestimmte Gameten[7] gebildet,
Gelb und bleich, liegen kleinere, männliche in ihrem Schlauch.
.
Nur eine heterothallische, Bryopsis plumosa[10], sprechen wir an,
Deren Weibchen und Männchen je ihre eignen Gametangien entwickeln;
Homothallische bilden beide Typen Gameten an ein und demselben siphonalen System.
.
Geißeleinziehend sich festsetzt und, mit außergewöhnlich großem
Zellkern versehen, mit angedeuteten Ästen leicht sich verzweigt,
Haploide[18] Kerne unüberschaubar vermehrt.
– Nur die Zygote ist demnach mit diploidem Zellkern bestückt; weil sie
Sofort die Meiose vollzieht, ist, was aus ihr wächst, kein Sporophyt[19]. –
.
Zwei Alternativen steh‘n der Zygote nun zur Verfügung:
Gametophyten als Weibchen und Männchen sich bilden,
Oder zu Stolonen zu wachsen mit beiden Gametophytengeschlechtern daran.
.
Gametangien werden wieder entstehen als septenbegrenzte Schläuche, mit
Zweigeißligen Schwärmern, kleine und große am vorausbestimmten Gezweig;
Finden sich, vereinen sich zur Zygote, wie davor schon Generationen:
Ihr Kreislauf des Lebens[22] geht wieder voran, wie‘s vor Tausenden Jahren schon war.
.
Fußnoten
[1] Stolonen (Bryopsidales): Siphonale, am Substrat entlangwachsende, oder im Sand verlaufende Schläuche, denen fertile strauch- oder bäumchenförmige Systeme entspringen
[2] Rhizoide: Fadenartige, wurzelähnliche, trichale oder unseptierte Auswüchse zum Festheften von Thallophyten
[3] Siphonale Röhren (Algen, Pilze): Vielkernige, lange Schläuche ohne untergliedernde Querwände
[4] Pinnat: Sind Elemente – wie Blättchen, Blattadern oder Äste – die auf beiden Seiten einer gemeinsamen Mittelachse, ähnlich wie die Strahlen an einer Feder, angeordnet sind.
[5] Gametangien: Behälter für Gameten; oder für Zellkerne, die, ohne Gameten zu bilden, dazu bestimmt sind, früher oder später über Karyogamie der sexuellen Fortpflanzung zu dienen
[6] Septum (Algen, Pilze): Querwand eines einzellreihigen Fadens, eines Trichoms, eines Zellausläufers, einer Hyphe
[7] Gameten: Für sexuelle Fortpflanzung vorgesehene haploide Zellen
[8] Homothallisch, monözisch (Algen): Weibliche und männliche Gameten werden auf dem gleichen Gametophyten gebildet
[9] Heterothallisch, diözisch (Algen): Weibliche und männliche Gameten werden auf unterschiedlichen Gametophyten gebildet
[10] Bryopsis plumosa: Flaumige Moosalge (Bryopsidaceae – Bryopsidales – Ulvophyceae – Chlorophyta – Plantae –…)
[11] Anisogamie (morphologische): Plasmo- und Karyogamie unterschiedlich gestalteter Gameten
[12] Plasmogamie: Verschmelzung der Protoplasten zweier Zellen im Zuge sexueller Fortpflanzung; abgekürzt P!
[13] Flagellum, Geißel (Eukaryageißel): Charakterisiert durch ihren internen Bau aus 9 peripheren, etwas schräg nach innen gestellten Doppelmikrotubuli (Querschnitt durch die Geißel) und durch ein zentrales Tubulipaar, das etwas Abstand voneinander hält. Dyneinarme verbinden die Mikrotubuli. Die Geißel ist von der Zellmembran umgeben und gefüllt mit Cytosol. Am Übergang der Geißelbasis in den Zellkörper treten spezielle Verstrebungen, Verstärkungen, auf; eine dünne Querplatte (Terminalplatte) trennt oft den untersten, in die Zelle integrierten Teil, der in seiner Struktur einem Centriol entspricht: Es fehlen die beiden zentralen Mikrotubuli und die peripheren Zwillinge wurden zu Drillingen. Die in der Zelle gelegenen Teile der Geißel sind noch durch verwandtschaftsabhängig gestaltete Haltestrukturen verwurzelt.
[14] Diploid: Zellkerne mit doppeltem Satz zusammenpassender, homologer, Chromosomen; ausgedrückt mit 2n
[15] Zygote: Diploide Zelle, die nach der Verschmelzung zweier haploider Kerne, im Zuge der sexuellen Fortpflanzung entstand
[16] Meiose, meiotisch, R!: Meiose dient der Reduktion eines diploiden Chromosomensatzes zu haploiden Sätzen. Dabei werden einander entsprechende Chromosomen, im Kern sich dann mittig in einer Ebene gegenüberstehend, gepaart und anschließend in entgegengesetzter Richtung („polwärts“) separiert. Dieser Vorgang wird auch als Reduktionsteilung (oft abgekürzt als R! und zugleich stellvertretend für die ganze Meiose verwendet) bezeichnet. Da die voneinander getrennten haploiden Chromosomen schon zu Chromatiden verdoppelt sind, schließt sich an die Reduktionsteilung noch eine mitotische Teilung an, so dass vier haploide Kerne letztlich vorliegen.
[17] Mitose, mitotisch, abgekürzt M!: Im Kern mittig in einer Ebene versammelte Chromosomen bildeten je zwei identische Chromatiden, die bei der Mitose durch Mikrotubuli separiert werden und, von einer Zellwand getrennt, als identische Chromosomensätze der entstandenen Zellen wirken
[18] Haploid: Zellkerne mit einfachem Chromosomensatz; ausgedrückt als n
[19] Sporophyt: Bildet im Generationswechsel von Sporophyt zu Gametophyt meiotisch (R!), da selbst diploid (oder dikaryotisch), haploide Sporen
[20] Sporangium: Behälter, in dem Sporen asexuell oder sexuell gebildet werden
[21] Stephanosporen: Zoosporen mit subapikalem Kranz aus Geißeln
[22] Lebenskreislauf, Lebenszyklus, Entwicklungszyklus, Entwicklungskreislauf: Ein Kreislauf (eigentlich ist es eine Schraube, weil immer Neues entsteht, das zwar dem Anfänglichen gleicht, doch zeitlich später kommt), in dem sexuelle Fortpflanzung erfolgt. Zusätzlich kann noch in regelmäßigem Wechsel eine Phase der asexuellen Vermehrung eingeschlossen sein.
Eingestellt am 21. Februar 2026
.

Oben: Bryopsis plumosa
{kind=link}
Autor: B.navez
Lizenz: GNU Free Documentation License; unverändert
Unten: Bryopsis vestita
{kind=link}
Autor: Peter de Lange
Lizenz: Public domain; unverändert
Eingestellt am 21. Februar 2026
.
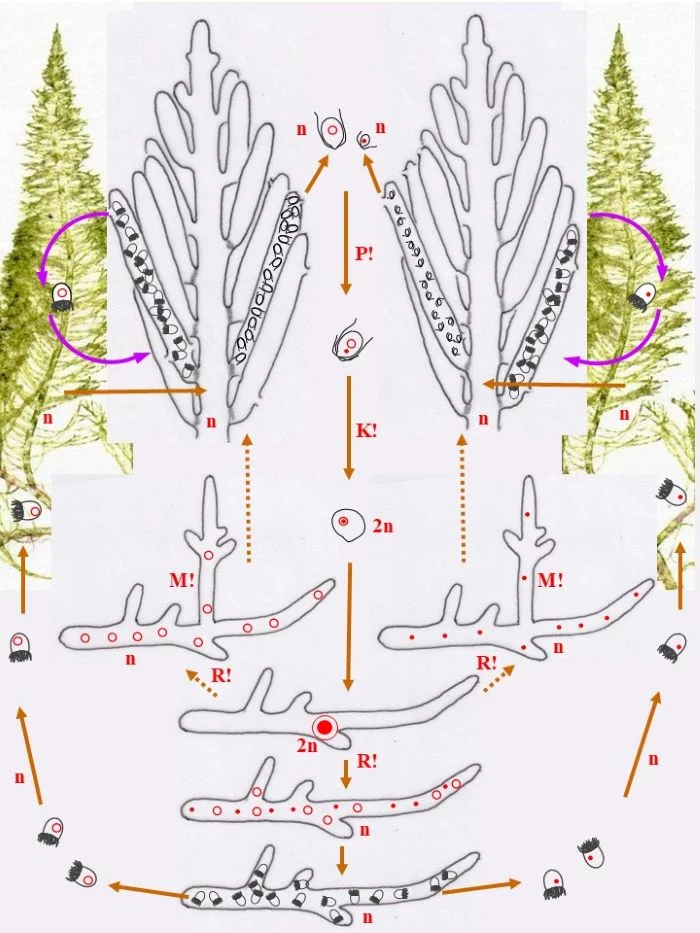
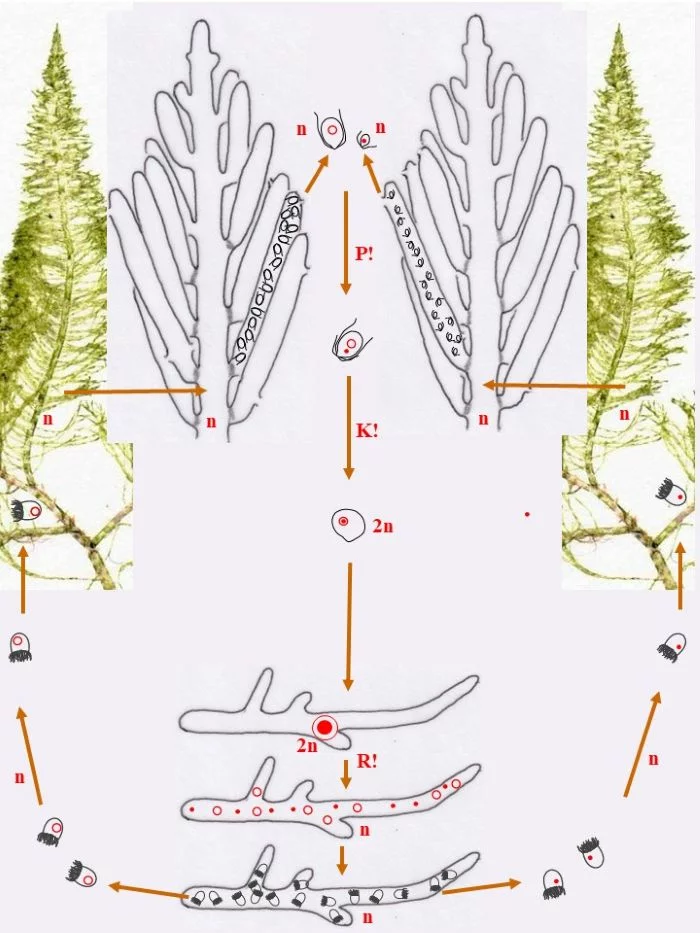
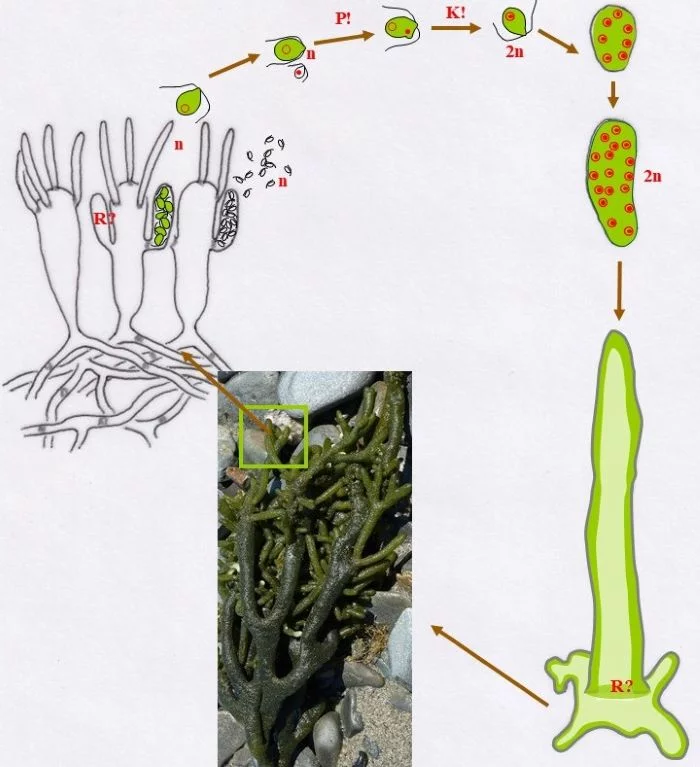
Entwicklungskreislauf von Bryopsis plumosa (Tusche- und ppt-generiert; Reinhard Agerer)
Die heterothallische, haploide (n) Flaumige Moosalge besteht aus zwei gleichgestalteten weiblichen und männlichen Gametophyten (Habitus, links und rechts). Federartig ist ihr siphonales Verzweigungssystem, dessen Endzweige im subterminalen Bereich weichliche und männliche Gametangien bildet, deren weibliche Gameten (groß; dargestellt mit Kringelkern) und männliche Gameten (klein; dargestellt mit Punktkern) durch Poren den Gametangien entschlüpfen. Die Gameten verschmelzen durch anisogame Plasmogamie (P!) und Karyogamie (K!) zur diploiden (2n) Zygote.
Die Zygote wächst zu einer diploiden, schlauchförmigen, kurz verzweigten Zygote aus, deren Kern (2n) zum Riesenzellkern (2n) wird; in ihr erfolgt die Meiose (R!) mit anschließenden Mitosen.
Mit den haploiden (n), männlichen (Punkt-Kerne) und weiblichen (Kringel-Kerne) Kernen entstehen haploide (n) Stephanosporen, die zu den entsprechenden Gametophyten auswachsen. Damit ist der sexuelle Entwicklungskreislauf geschlossen (durchgezogene braune Pfeile), aber
Bryopsis plumosa besitzt noch eine abweichende Möglichkeit, sich zu den Gametophyten zu entwickeln (gepunktete braune Pfeile): Die Zygote wächst unter Meiose (R!) gleich zu weiblichen und männlichen Gametophyten aus, deren Zellkerne offensichtlich sortiert (auf welche Weise?) in die Gametophyten gelangen. Mit dieser stephanosporenlosen Variante ist ebenso der sexuelle Entwicklungskreislauf geschlossen.
Auch asexuelle Vermehrung der Gametophyten ist der Flaumigen Moosalge möglich: Anstelle von Gametangien entstehen Sporangien mit Stephanosporen, die freiwerdend,
Nebenkreisläufe initiieren (violette Pfeile), wodurch aus den Stephanosporen neue, genetisch identische Gametophyten entstehen. Es liegt also ein haplontischer Entwicklungszyklus vor, bei dem nur die Zygote diploid ist, die allerdings einen Riesenzellkern entwickelt. Nur die Zygote ist diploid (2n), somit liegt ein zygotischer Kernphasenwechsel vor.
Nach van den Hoek et al. (1995/2002), Fig. 24.7, S. 427.
In den nachfolgenden sechs Abbildungen ist der Entwicklungszyklus einer Powerpoint-Präsentation ähnlich aufgebaut.
Eingestellt am 21. Februar 2026
.
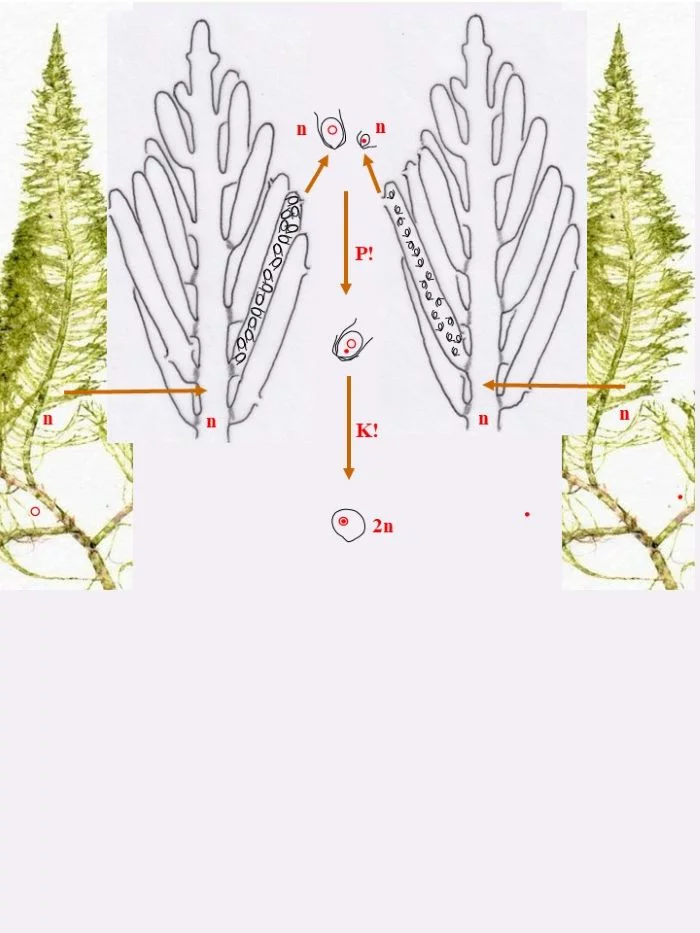
Die heterothallische, haploide (n) Flaumige Moosalge besteht aus zwei gleichgestalteten weiblichen und männlichen Gametophyten (Habitus, links und rechts). Federartig ist ihr siphonales Verzweigungssystem, dessen Endzweige im subterminalen Bereich weichliche und männliche Gametangien bildet, deren weibliche Gameten (groß; dargestellt mit Kringelkern) und männliche Gameten (klein; dargestellt mit Punktkern) durch Poren den Gametangien entschlüpfen. Die Gameten verschmelzen durch anisogame Plasmogamie (P!) und Karyogamie (K!) zur diploiden (2n) Zygote.
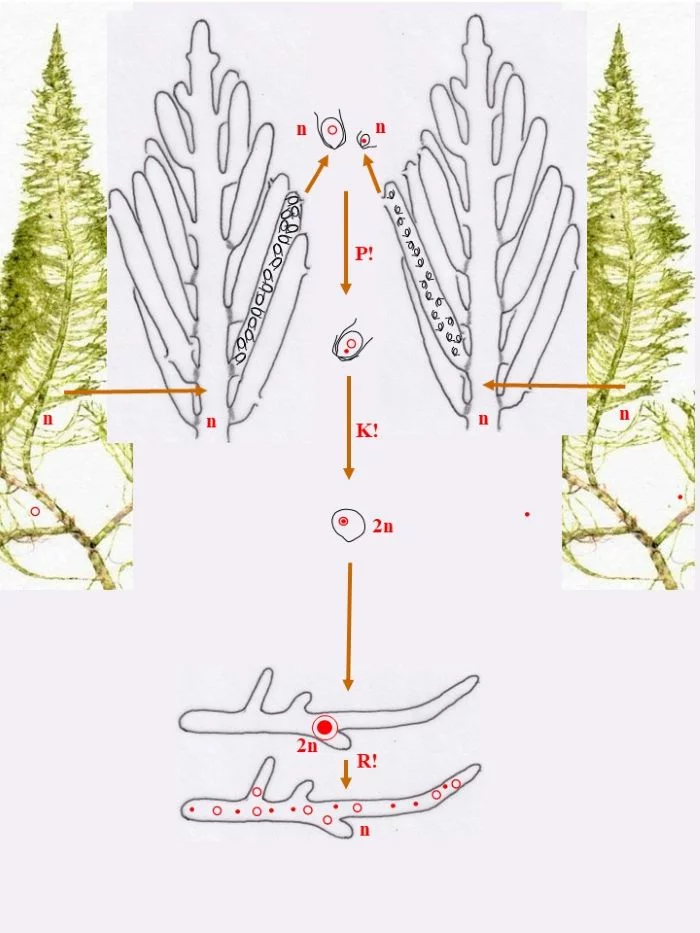
Die Zygote wächst zu einer diploiden, schlauchförmigen, kurz verzweigten Zygote aus, deren Kern (2n) zum Riesenzellkern (2n) wird; in ihr erfolgt die Meiose (R!) mit anschließenden Mitosen.
Mit den haploiden (n), männlichen (Punkt-Kerne) und weiblichen (Kringel-Kerne) Kernen entstehen haploide (n) Stephanosporen, die zu den entsprechenden Gametophyten auswachsen. Damit ist der sexuelle Entwicklungskreislauf geschlossen (durchgezogene braune Pfeile), aber
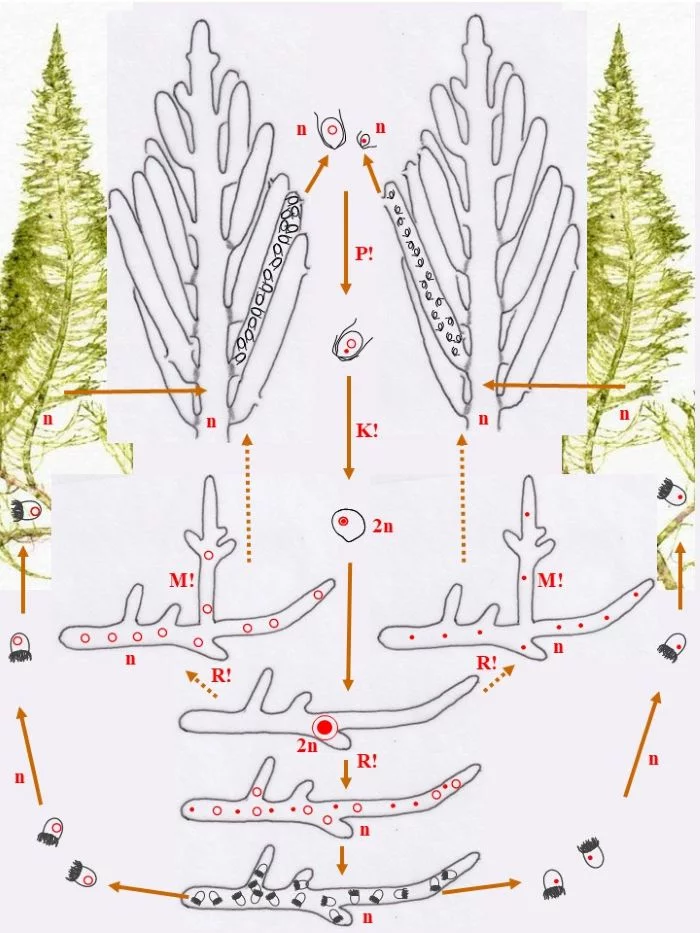
Bryopsis plumosa besitzt noch eine abweichende Möglichkeit, sich zu den Gametophyten zu entwickeln (gepunktete braune Pfeile): Die Zygote wächst unter Meiose (R!) gleich zu weiblichen und männlichen Gametophyten aus, deren Zellkerne offensichtlich sortiert (auf welche Weise?) in die Gametophyten gelangen. Mit dieser stephanosporenlosen Variante ist ebenso der sexuelle Entwicklungskreislauf geschlossen.
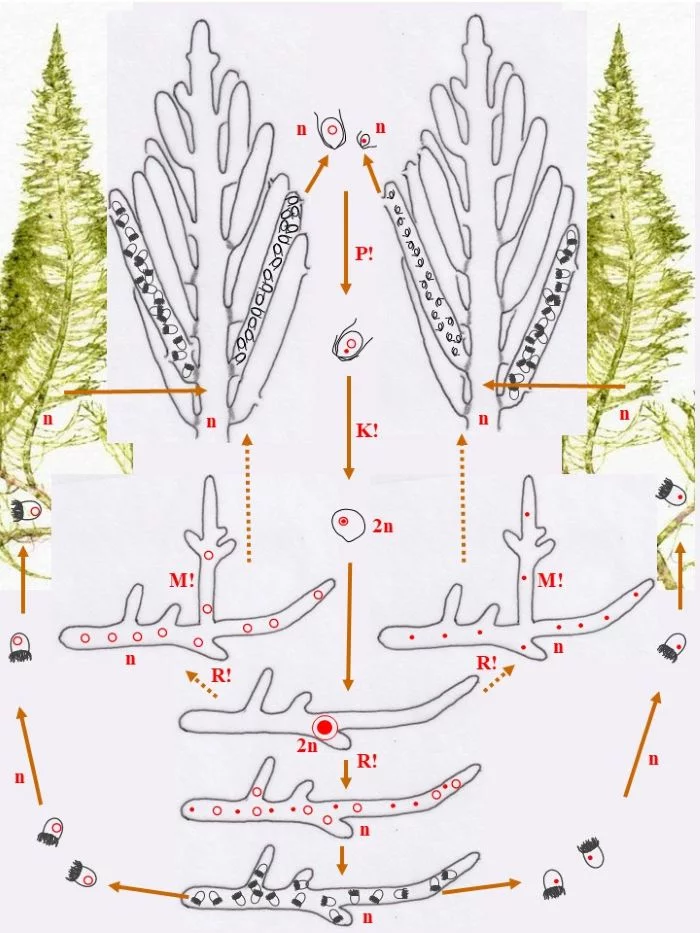
Auch asexuelle Vermehrung der Gametophyten ist der Flaumigen Moosalge möglich: Anstelle von Gametangien entstehen Sporangien mit Stephanosporen, die freiwerdend,
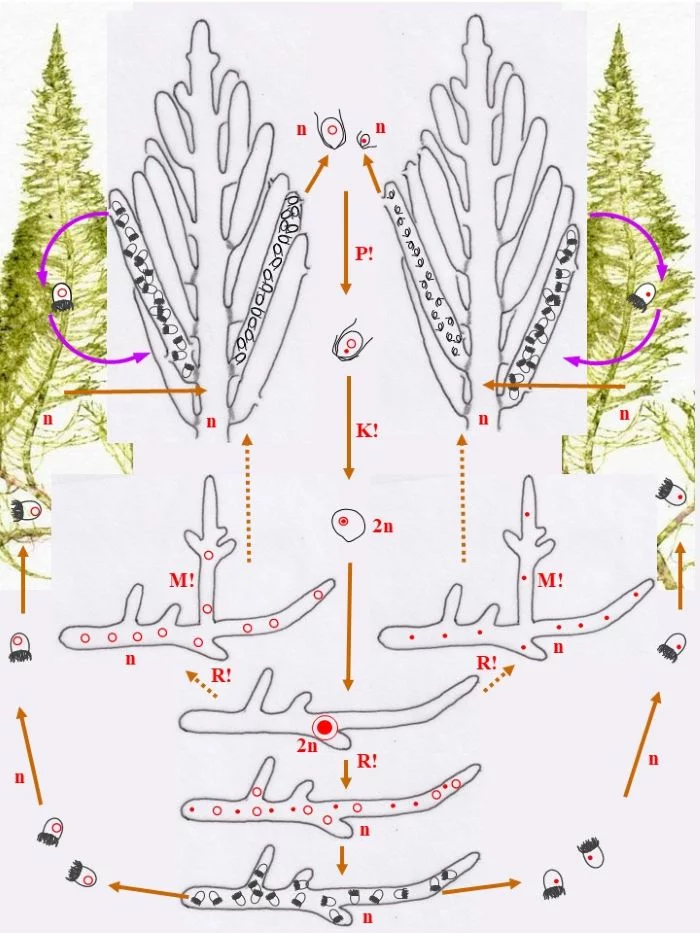
Nebenkreisläufe initiieren (violette Pfeile), wodurch aus den Stephanosporen neue, genetisch identische Gametophyten entstehen. Es liegt also ein haplontischer Entwicklungszyklus vor, bei dem nur die Zygote diploid ist, die allerdings einen Riesenzellkern entwickelt. Nur die Zygote ist diploid (2n), somit liegt ein zygotischer Kernphasenwechsel vor.
Nach van den Hoek et al. (1995/2002), Fig. 24.7, S. 427.
Eingestellt am 21. Februar 2026
.
Bryopsidaceae, Moosalgen i.e.S.
2 Blasiger Diplo-Haplont?
.
Grüne Finger liegen am Strand, verzweigt, aneinanderhängend wie an Gelenken,
Eines Toten Manns Finger[1] – die mit Algen grün überlaufen? – wie die Moosalge heißt.
Oder einfach nur Meer-Hirschgeweih? Auch dies scheint mit Algen grün überzogen.
Ohne menschliche Assoziation heißt sie Fragile[2], zerbricht sie doch an ihren Taillen recht leicht.
.
Ein Teil der Pflanze ist es nur, die jemand am Sandstrand gefunden.
Vielleicht war sie, wie die längsten so werden, gar einen Meter und darüber noch lang.
Niemand hat noch gemessen – wer schon würde dies wollen? –
Wie lang der Schlauch[3], der diese Alge gebaut.
.
Ein Schnitt mit dem Messer, mit der Rasierklinge, sie zu mikroskopieren,
Zeigt eine grünliche Masse, die dem Fund entquillt:
Kam aus zerschnitt’nen, zerbroch’nen, schottenlosen[6] Schläuchen hervor.
Stabilisiert werden die Röhren durch wandständige Ringe,
Doch, um nach Bruch einer Zelle, größeren Protoplastenverlust zu verhindern,
Werden Schläuche vielerorts mit Pfropf – keine Querwand! – unterteilt.
.
Außen herum liegen dicht dicke, längsgezogene Blasen,
Aus denen so manche kleinere subapikal oder seitlich trieb.
Im Zentrum aber der Fingerglieder, das Experten Medulla[10] nennen,
Liegen ungeordnete Schläuche längs, kreuz und quer,
Nur gegen die Blasen erscheinen sie dichter und eher geregelt,
Um Blasen ohne Lücken zu stellen, sie zu versammeln als durchgehende Schicht.
.
Blasen zweiter Ordnung, die kleineren, aus den großen getrieben,
Sind Gametangien[11], vom Träger je durch ein Septum begrenzt,
Männliche und weibliche steh’n durcheinander, gleiche wohl aber beisammen,
Was der großen Blase, Träger der kleinen, entspringt.
Überragt werden die großen von dünneren, kürzeren Schläuchen,
Die haarförmig und Gametangienschutz wohl sind.
.
Männliche, gelbliche Gametangien, teilen den Inhalt in
Winzige biflagellate männliche Gameten[12] auf;
Weibliche Gametangien sind, wegen der vielen Chloroplasten in
Mehr als zehnmal so großen, ebenfalls biflagellaten Gameten, dunkelgrün.
.
Die zweigeißlig[17], wie der weibliche Gamet, geblieben, eine Weile noch schwimmt,
Bis sie sich, Untergrund kontaktierend, Geißeln verlierend, festheftet
Und zur ersten Schlauchgestalt wächst. –
.
Was der Wissenschaft noch nicht endgültig gelungen[18], ist,
Festzustellen, wo die Meiose[19] erfolgt:
Gleich nach Karyogamie zur Zygote,
Bevor sie zur Schlauchgestalt wächst,
Damit ein haploider Gametophyt so entsteht,
Somit Gameten nur mitotisch sich bilden;
Oder, dass der Kern der Zygote mitotisch sich teilt
Und folglich ein diploider Sporophyt entsteht,
Wonach an ihm erst in Gametangien
Meiotische Teilung erfolgt.
.
Im ersten Fall würde ein haplontischer[22] Kreislauf bestehen,
Bei dem die Zygote nur doppelten Chromosomensatz hat,
Im zweiten ein diplontischer[23], bei dem Gameten in des Sporophyten Gametangien
Erst nach Meiose entsteh‘n und so nur sie haploid sind.
.
Werden jedoch anderer Bryopsidaceen Lebenskreisläufe[24] betrachtet und mit
Dem von Codium verglichen, lässt der Schluss sich ohne Weiteres zieh’n:
Das Meergeweih repräsentiert den Sporophyten,
Denn all anderer Arten Sporophyten führen Meiose durch,
Um haploide Stephanosporen zu bilden,
Für männlicher und weiblicher Gametophyten Start.
.
Codium, so lässt sich folgern, reduzierte Gametophyten,
Sparte damit haploide Stephanosporen ebenfalls ein
Und verlegte Meiosen zum Sporophyten,
Der nun in Gametangien nach Meiosen haploide Gameten entstehen lässt.
.
Fußnoten
[1] Toter-Manns-Finger, Meergeweih: Codium fragile (Bryopsidaceae – Bryopsidales – Ulvophyceae – Chlorophyta – Plantae – …)
[2] Codium fragile: Toten-Manns-Finger, Meergeweih (Bryopsidaceae – Bryopsidales – Ulvophyceae – Chlorophyta – Plantae – …)
[3] Siphonal gebaut
[4] Protoplast, Protoplasma: Gesamter Inhalt einer Zelle
[5] Chloroplast (allgemein): Zur Fotosynthese befähigter, grüner Chromatophor
[6] Septenlos, querwandlos
[7] Mannan: Polymere aus β-1,4 glycosidisch verknüpfen Mannose-Einfachzucker
[8] Sulfatiert: mit Sulfat [SO4–] verbunden
[9] Arabinoglucan: heterogenes Polysaccharid aus Arabinose und Glucose
[10] Medulla (Laminariales, Bryopsidales): Aufgelockertes, matrixerfülltes Zentrum, von hyphenartigen Fäden oder von siphonalen Schläuchen durchzogen
[11] Gametangien: Behälter für Gameten; oder für Zellkerne, die, ohne Gameten zu bilden, dazu bestimmt sind, früher oder später über Karyogamie der sexuellen Fortpflanzung zu dienen
[12] Gameten: Für sexuelle Fortpflanzung vorgesehene haploide Zellen
[13] Plasmogamie: Verschmelzung der Protoplasten zweier Zellen im Zuge sexueller Fortpflanzung; abgekürzt P!
[14] Karyogamie: Verschmelzung zweier haploider Zellkerne; abgekürzt K!
[15] Diploid: Zellkerne mit doppeltem Satz zusammenpassender, homologer, Chromosomen; ausgedrückt mit 2n
[16] Zygote: Diploide Zelle, die nach der Verschmelzung zweier haploider Kerne, im Zuge der sexuellen Fortpflanzung entstand
[17] Flagellum, Geißel (Eukaryageißel): Charakterisiert durch ihren internen Bau aus 9 peripheren, etwas schräg nach innen gestellten Doppelmikrotubuli (Querschnitt durch die Geißel) und durch ein zentrales Tubulipaar, das etwas Abstand voneinander hält. Dyneinarme verbinden die Mikrotubuli. Die Geißel ist von der Zellmembran umgeben und gefüllt mit Cytosol. Am Übergang der Geißelbasis in den Zellkörper treten spezielle Verstrebungen, Verstärkungen, auf; eine dünne Querplatte (Terminalplatte) trennt oft den untersten, in die Zelle integrierten Teil, der in seiner Struktur einem Centriol entspricht: Es fehlen die beiden zentralen Mikrotubuli und die peripheren Zwillinge wurden zu Drillingen. Die in der Zelle gelegenen Teile der Geißel sind noch durch verwandtschaftsabhängig gestaltete Haltestrukturen verwurzelt
[18] Dies ist der großen Kernzahl bei siphonaler Organisation geschuldet; erst Messungen von DNA-Gehalten von Kernen könnte die Frage lösen; Meiosen der Zygote wurden offenbar noch nicht direkt beobachtet.
[19] Meiose, meiotisch, R!: Meiose dient der Reduktion eines diploiden Chromosomensatzes zu haploiden Sätzen. Dabei werden einander entsprechende Chromosomen, im Kern sich dann mittig in einer Ebene gegenüberstehend, gepaart und anschließend in entgegengesetzter Richtung („polwärts“) separiert. Dieser Vorgang wird auch als Reduktionsteilung (oft abgekürzt als R! und zugleich stellvertretend für die ganze Meiose verwendet) bezeichnet. Da die voneinander getrennten haploiden Chromosomen schon zu Chromatiden verdoppelt sind, schließt sich an die Reduktionsteilung noch eine mitotische Teilung an, so dass vier haploide Kerne letztlich vorliegen.
[20] Haploid: Zellkerne mit einfachem Chromosomensatz; ausgedrückt als n
[21] Mitose, mitotisch, abgekürzt M!: Im Kern mittig in einer Ebene versammelte Chromosomen bildeten je zwei identische Chromatiden, die bei der Mitose durch Mikrotubuli separiert werden und, von einer Zellwand getrennt, als identische Chromosomensätze der entstandenen Zellen wirken
[22] Haplontischer Entwicklungszyklus: Bei diesem Lebenskreislauf ist ausschließlich die Zygote diploid, alles andere haploid
[23] Diplontischer Entwicklungszklus; Bei diesem Lebenskreislauf sind ausschließlich Gameten haploid.
[24] Lebenskreislauf, Lebenszyklus, Entwicklungszyklus, Entwicklungskreislauf: Ein Kreislauf (eigentlich ist es eine Schraube, weil immer Neues entsteht, das zwar dem Anfänglichen gleicht, doch zeitlich später kommt), in dem sexuelle Fortpflanzung erfolgt. Zusätzlich kann noch in regelmäßigem Wechsel eine Phase der asexuellen Vermehrung eingeschlossen sein.
Eingestellt am 21. Februar 2026
.

Codium fragile (Toten-Manns-Finger, Meergeweih)
_-_Tor_Bay_Provincial_Park,_Nova_Scotia_2022-07-27_(01).jpg){kind=link}
Autor: Ryan Hodnett
Lizenz: Creative Commons Attribution-Share Alike 4.0 International license; unverändert
Eingestellt am 21. Februar 2026
.
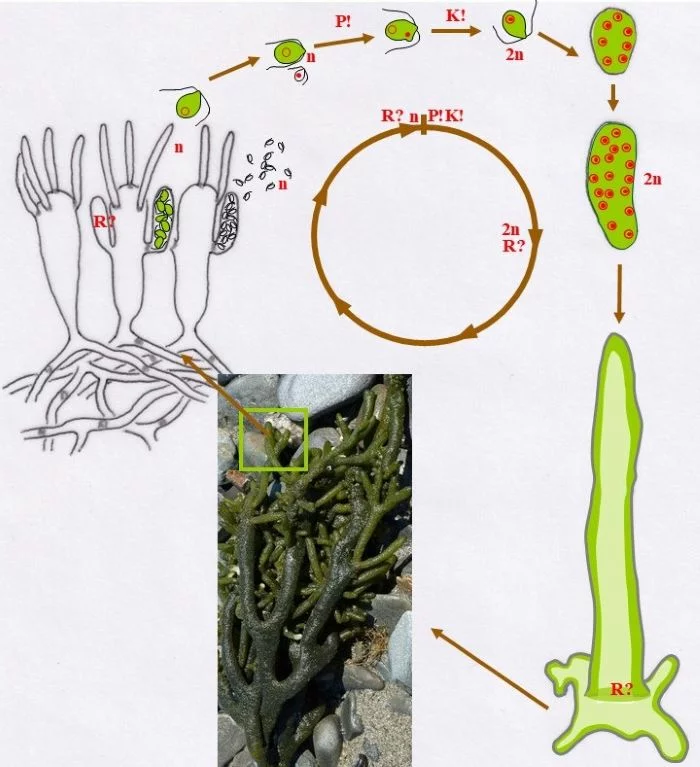
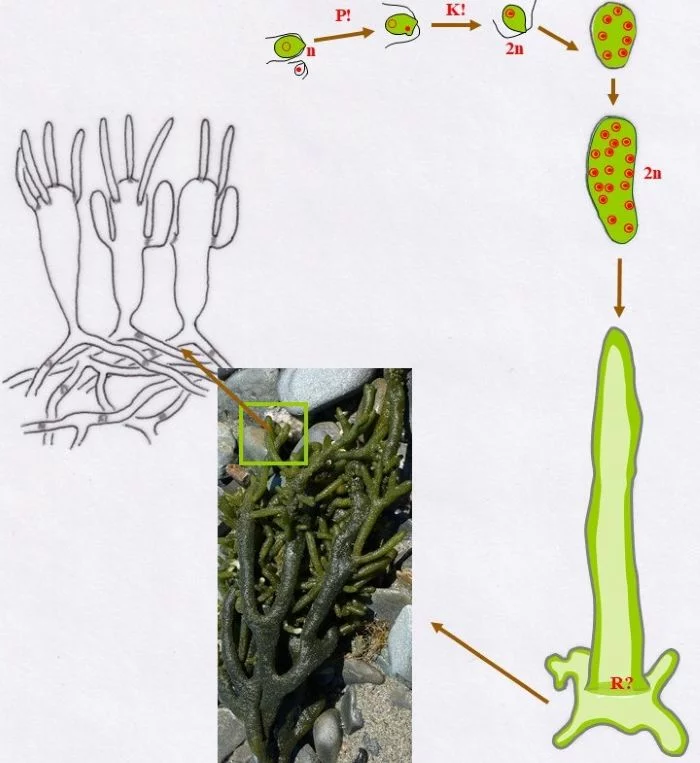
Entwicklungszyklus von Codium fragile
Unter Anisogamie verschmelzen (P!) ein großer, chloroplastenreicher, weiblicher Gamet (Kringel-Kern) und ein sehr kleiner, chloroplastenloser männlicher Gamet (Punkt-Kern) und vollziehen danach die Karyogamie (K!) zur diploiden (2n) Zygote.
Die Zygote setzt sich fest auf einem Substrat, teilt mitotisch die Kerne, so dass ein vielkerniges, diploides, junges Codium fragile entsteht. Danach wächst der Keimling zu einem vielkernigen Schlauch aus, heftet sich mit Auswüchsen an das Substrat breitflächig an, vakuolisiert zentral und schiebt damit den Protoplasten mit Zellkernen und Chloroplasten an die Peripherie (dunkler grün).
Durch vielfache Verzweigungen des siphonalen Schlauchs, entsteht mit der Zeit ein größeres Pflänzchen, das aus einem Plectenchym besteht. Zentral liegen die Schläuche lockerer, besitzen auch weniger Chloroplasten, randlich verdichten sie sich, an denen keulige Blasen („Vesikel“) entstehen, die ihrerseits nach außen kurze, haarartige Zellen bilden. Das siphonale System wird in unterschiedlichen Abständen durch Pfropfen (keine Querwände) untergliedert.
An diesen Blasen entstehen seitliche, sich nach oben entwickelnde Auswüchse als Gametangien, in denen sich je nach Blase, von der sie gebildet werden, große, grüne, weibliche Gameten entwickeln oder viele kleine, blasse männliche. Die Gameten sind definitionsgemäß haploid. – Wo Meiose (R?) stattfindet, ist offenbar noch unklar und wegen der vielen Zellkerne auch schwer zu bestimmen. Sie müsste entweder beim Auskeimen der Zygote zum Schlauch stattfinden (damit müssten alle Kerne die Meiose vollziehen; oder nur ein Teil, die anderen aber absterben) oder vor der Gametenbildung. Doch, um nur geschlechtsidentische Gameten im Gametangien entstehen zu lassen, dürfte nur ein sich mehrfach mitotisch teilender haploider Kern ins künftige Gametangium gelangen oder ein diploider, von dem drei der vier haploiden Kerne zugrunde gehen, der übrige aber sich mehrfach mitotisch teilt. Zu überlegen wäre auch ein anderer eleganterer Weg: Die Meiose mit drei absterbenden haploiden Kernen findet bereits im „Vesikel“ statt; mitotisch vermehrte gleichgeschlechtliche Kerne können dann wahllos in die zukünftigen Gametangien eintreten und Gameten bilden; damit würden auch alle sich aus einer Blase entwickelnden Gametangien Gameten gleichen Geschlechts besitzen. –
In Kreisform dargestellt, zeigen sich nur drei Eigenschaften als entschieden: (a) In Gametangien werden haploide Gameten gebildet, die (b) über Plasmogamie und Karyogamie eine diploide Zygote entstehen lassen und (c) die sich, vielkernig werdend, irgendwo festsetzt und zumindest eine noch anfangs diploide Pflanze bildet.
Nach van den Hoek et al. (1995/2002), Fig. 24.10, S. 431.
In den nachfolgenden fünf Abbildungen ist der Entwicklungszyklus einer Powerpoint-Präsentation ähnlich aufgebaut
Eingestellt am 21. Februar 2026
.
Unter Anisogamie verschmelzen (P!) ein großer, chloroplastenreicher, weiblicher Gamet (Kringel-Kern) und ein sehr kleiner, chloroplastenloser männlicher Gamet (Punkt-Kern) und vollziehen danach die Karyogamie (K!) zur diploiden (2n) Zygote.
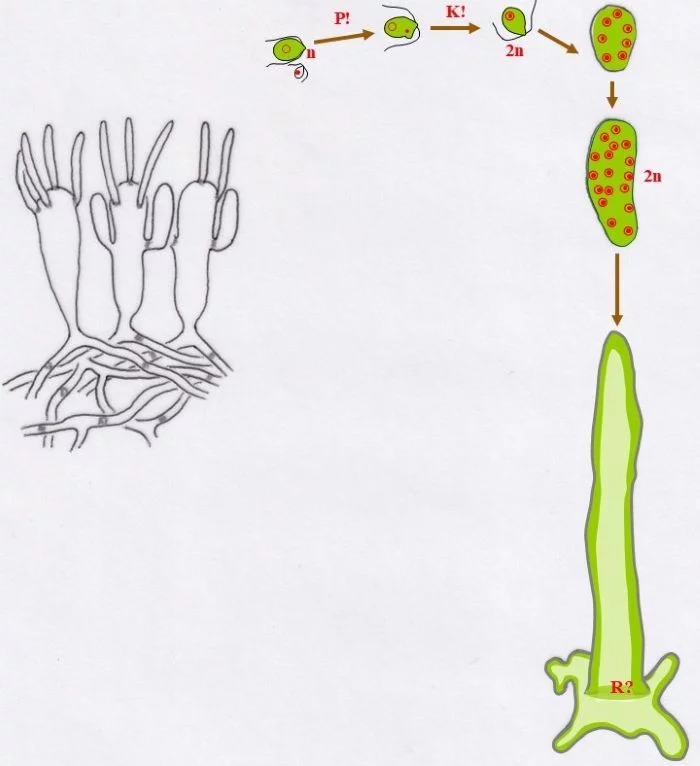
Die Zygote setzt sich fest auf einem Substrat, teilt mitotisch die Kerne, so dass ein vielkerniges, diploides, junges Codium fragile entsteht. Danach wächst der Keimling zu einem vielkernigen Schlauch aus, heftet sich mit Auswüchsen an das Substrat breitflächig an, vakuolisiert zentral und schiebt damit den Protoplasten mit Zellkernen und Chloroplasten an die Peripherie (dunkler grün).
Durch vielfache Verzweigungen des siphonalen Schlauchs, entsteht mit der Zeit ein größeres Pflänzchen, das aus einem Plectenchym besteht. Zentral liegen die Schläuche lockerer, besitzen auch weniger Chloroplasten, randlich verdichten sie sich, an denen keulige Blasen („Vesikel“) entstehen, die ihrerseits nach außen kurze, haarartige Zellen bilden. Das siphonale System wird in unterschiedlichen Abständen durch Pfropfen (keine Querwände) untergliedert.
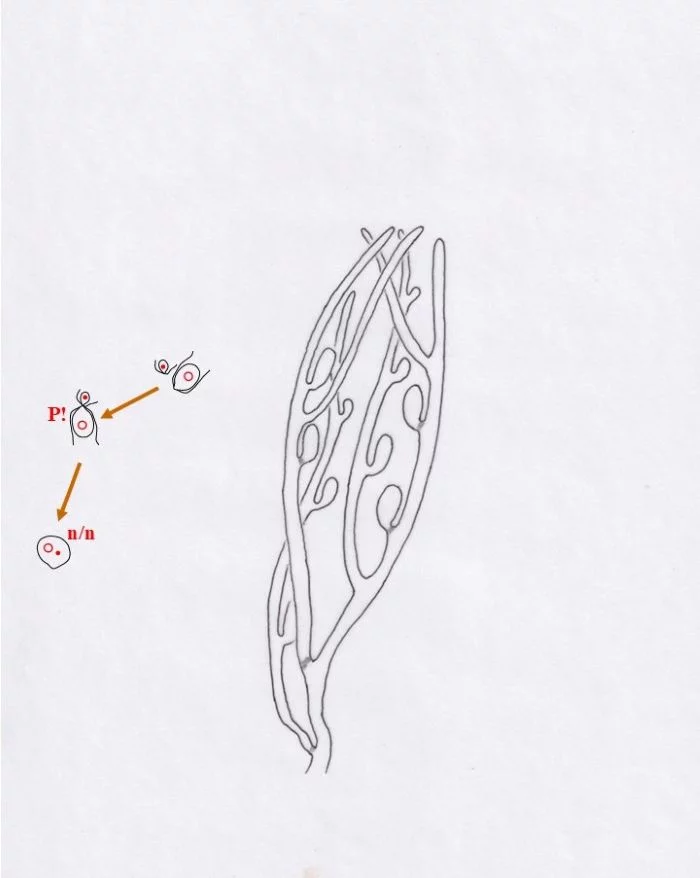
An diesen Blasen entstehen seitliche, sich nach oben entwickelnde Auswüchse als Gametangien, in denen sich je nach Blase, von der sie gebildet werden, große, grüne, weibliche Gameten entwickeln oder viele kleine, blasse männliche. Die Gameten sind definitionsgemäß haploid. – Wo Meiose (R?) stattfindet, ist offenbar noch unklar und wegen der vielen Zellkerne auch schwer zu bestimmen. Sie müsste entweder beim Auskeimen der Zygote zum Schlauch stattfinden (damit müssten alle Kerne die Meiose vollziehen; oder nur ein Teil, die anderen aber absterben) oder vor der Gametenbildung. Doch, um nur geschlechtsidentische Gameten im Gametangien entstehen zu lassen, dürfte nur ein sich mehrfach mitotisch teilender haploider Kern ins künftige Gametangium gelangen oder ein diploider, von dem drei der vier haploiden Kerne zugrunde gehen, der übrige aber sich mehrfach mitotisch teilt. Zu überlegen wäre auch ein anderer eleganterer Weg: Die Meiose mit drei absterbenden haploiden Kernen findet bereits im „Vesikel“ statt; mitotisch vermehrte gleichgeschlechtliche Kerne können dann wahllos in die zukünftigen Gametangien eintreten und Gameten bilden; damit würden auch alle sich aus einer Blase entwickelnden Gametangien Gameten gleichen Geschlechts besitzen. –
In Kreisform dargestellt, zeigen sich nur drei Eigenschaften als entschieden: (a) In Gametangien werden haploide Gameten gebildet, die (b) über Plasmogamie und Karyogamie eine diploide Zygote entstehen lassen und (c) die sich, vielkernig werdend, irgendwo festsetzt und zumindest eine noch anfangs diploide Pflanze bildet.
Nach van den Hoek et al. (1995/2002), Fig. 24.10, S. 431.
Eingestellt am 21. Februar 2026
.
Bryopsidaceae, Moosalgen i.e.S.
4 Blasiger Dikaryo-Haplont
.
Lange rätselten Experten über Halicystis olvalis‘ vollständigen Lebenskreislauf[1],
Fanden sie doch immer zweigeißlige[2] Flagellaten, groß und klein, aus zwei
Unterschiedlichen, doch gleichgestalteten halbcentimetergroßen kugelförmigen Blasen
Durch eine oder mehrere Poren treten; beobachtete vielleicht auch schon Anisogamie[3].
Jene der größeren Schwärmer dagegen dunkelgrüne nahe der Wand.
Verankert waren die einzelligen, runden Behälter jeweils mit
Ausgezogenem Stielchen, nicht verschwemmt zu werden, im Substrat[6]. –
.
Zu einer anderen Zeit fanden und analysierten Phykologen[7] eine
In oft dichter Versammlung große, eiförmige Behälter trugen, die viele Kerne[10] enthielten
Und, wenn einer älter geworden, Stephanosporen[11] in die Umwelt gab, die,
Irgendwo sich niederließen für etwas, was der Entdeckung noch harrte.
Neun Jahrzehnte freilich verrannen, bis die Wissenschaft das aufeinander Angewiesensein
.
Diese Entdeckung hatte zur Folge, dass für beide Generationen
Derbesia marina[16] nun der gültige Name war,
Wurde Derbesia marina doch um sechs Jahre früher beschrieben als Halicystis ovalis,
Denn für einen Organismus, der in zwei Generationen lebt, ist nur der früher verliehene Name erlaubt, hat deshalb Priorität[17].
.
Die Fragen gingen aber noch weiter:
Festzustellen galt, was mit den Kernen geschieht:
Wann und wo Plasmogamie[18], Karyogamie und Meiose erfolgen?
Oder besitzen sie gar einen Diplo-haplontischen Entwicklungszyklus[21]?
Die Überraschung gelang Derbesia marina perfekt:
.
Durch Anisogamie verschmelzen die ungleichgroßen Zweigeißelgameten.
Der kleinere, der männliche, übergibt dem weiblichen nur seinen Kern,
Er selbst bleibt außerhalb, verschwindet ganz leise; der weibliche lässt die Beiden zusammen,
Vereint jedoch sie nicht und wächst zum siphonalen System, zum Sporophyten[22] heran!
Nur am Ende, wenn es Stephanosporen in Derbesias Blasen zu bilden gilt,
Paaren sich viele der männlichen und weiblichen Kerne im Apex der Blasen.
Die anderen scheinen davon vollkommen unberührt;
Sie versammeln verschämt sich am unteren Ende;
.
Die, freigesetzt aus der zum Sporangium[30] gewordenen Blase, fortschwimmen, bis sie
Irgendwo landen, sich selbst zur Blase mit Haltestielchen entwickeln und
Die Gametangien, je nach der Gameten Chloroplastengehalt, unterschiedliche Tönung geben:
Gelbliche männlichen Gametangien, grüne weiblichen, nehmen doch größere Gameten mehr Chloroplasten mit.
.
Zumindest in des späten Sporangiums oberem Teil;
Kerne erscheinen dort miteinander weitgehend gekoppelt, wie in dikaryotischen Fungi[35];
Im siphonalen Röhrensystem liegen sie, womöglich aneinander uninteressiert und
durchmischt. –
.
Am Ende bleibt noch ein wichtiges Detail zu erwähnen,
Das hilft, auch andere Entwicklungszyklen der Bryopsidaceen gut zu versteh’n:
.
Im Gametophyten von Bryopsis herrscht mit etwas Cellulose Xylan weitgehend vor,
In der Zygote und im angedeutet verzweigten, noch gering entwickelten
Diploiden Sporophyten hingegen Mannan.
.
Codium, bei dem die Frage bestand, ob ein haploider oder ein diploider Thallus[44] vorliegt,
Besitzt als strukturelles Polysaccharid hauptsächlich Mannan
Und dürfte daher den diploiden Sporophyten repräsentieren, somit läge bei
Codium ein gametischer Kernphasenwechsel[45] vor, wobei nur die Gameten haploid sind. –
.
Fußnoten
[1] Lebenskreislauf, Lebenszyklus, Entwicklungszyklus, Entwicklungskreislauf: Ein Kreislauf (eigentlich ist es eine Schraube, weil immer Neues entsteht, das zwar dem Anfänglichen gleicht, doch zeitlich später kommt), in dem sexuelle Fortpflanzung erfolgt. Zusätzlich kann noch in regelmäßigem Wechsel eine Phase der asexuellen Vermehrung eingeschlossen sein.
[2] Flagellum, Geißel (Eukaryageißel): Charakterisiert durch ihren internen Bau aus 9 peripheren, etwas schräg nach innen gestellten Doppelmikrotubuli (Querschnitt durch die Geißel) und durch ein zentrales Tubulipaar, das etwas Abstand voneinander hält. Dyneinarme verbinden die Mikrotubuli. Die Geißel ist von der Zellmembran umgeben und gefüllt mit Cytosol. Am Übergang der Geißelbasis in den Zellkörper treten spezielle Verstrebungen, Verstärkungen, auf; eine dünne Querplatte (Terminalplatte) trennt oft den untersten, in die Zelle integrierten Teil, der in seiner Struktur einem Centriol entspricht: Es fehlen die beiden zentralen Mikrotubuli und die peripheren Zwillinge wurden zu Drillingen. Die in der Zelle gelegenen Teile der Geißel sind noch durch verwandtschaftsabhängig gestaltete Haltestrukturen verwurzelt.
[3] Anisogamie (morphologische): Plasmo- und Karyogamie unterschiedlich gestalteter Gameten
[4] Schwärmer, Zoid: Allgemeiner Ausdruck für begeißelte, bewegliche Zellen (Zoospore oder Gamet)
[5] Parietal: an der Wand, wandständig, der Wand entlang
[6] Substrat: Allgemeine Bezeichnung für feste, unterstützende, tragende oder nährende Substanz
[7] Phykologen: Sich mit Algen beschäftigende Experten
[8] Siphonal (Algen, Pilze): Vielkernige, lange Schläuche ohne untergliedernde Querwände
[9] Derbesia marina, als Art 1846 beschrieben
[10] Zellkern
[11] Stephanosporen: Zoosporen mit subapikalem Kranz aus Geißeln
[12] Halicystis ovalis, als Art 1850 beschrieben
[13] Publizert im Jahre 1938
[14] Haplont: Organismus, dessen Zygote ausschließlich diploid ist, alles andere ist haploid
[15] Diplont: Organismus, bei dem ausschließlich die Gameten haploid sind, alles andere ist diploid
[16] Derbesia marina: Meerbüschelalge (Bryopsidaceae – Bryopsidales – Ulvophyceae – Chlorophyta – Plantae –…)
[17] Priorität (Taxonomie): Beim Benennen einer Art gilt nach dem International Code of Botanical Nomenclature (ICBN) jener Name als einzig gültig, falls dem gleichen Organismus unterschiedliche gegeben wurden, dessen Publikationsdatum das älteste ist und der Name gültig (u. a. als Binomen; in einem zugänglichen Publikationsorgan; mit einer aussagekräftigen Beschreibung) publiziert wurde.
[18] Plasmogamie: Verschmelzung der Protoplasten zweier Zellen im Zuge sexueller Fortpflanzung; abgekürzt P!
[19] Haplontischer Entwicklungszyklus: Bei diesem Lebenskreislauf ist ausschließlich die Zygote diploid, alles andere haploid
[20] Diplontischer Entwicklungszklus; Bei diesem Lebenskreislauf sind ausschließlich Gameten haploid
[21] Diplo-haplontischer Entwicklungszyklus: Hier wechseln sich diploide, auf Grund einer Befruchtung (B! oder getrennt ausgedrückt: P! + K!) entstandene und haploide, auf Grund von Meiose (R!) gebildete Generationen regelmäßig ab.
[22] Sporophyt: Bildet im Generationswechsel von Sporophyt zu Gametophyt meiotisch (R!), da selbst diploid (oder dikaryotisch), haploide Sporen.
[23] Mitose, mitotisch, abgekürzt M!: Im Kern mittig in einer Ebene versammelte Chromosomen bildeten je zwei identische Chromatiden, die bei der Mitose durch Mikrotubuli separiert werden und, von einer Zellwand getrennt, als identische Chromosomensätze der entstandenen Zellen wirken.
[24] Heterokaryotisch: Zelle, die zwei oder mehr genetisch unterschiedliche Zellkerne in einem gemeinsamen Zytoplasma enthält; im Extremfall männliche und weibliche Kerne, die (noch) kein Dikaryon bilden.
[25] Dikaryon, dikaryotisch: Zwei konträrgeschlechtliche haploide (n) Kerne [werden als (+)- und (–)-Kerne oder als α- und β-Kerne bezeichnet; unterscheiden sich ihre Behälter in Größe oder die Dikaryen bildenden Kerne in der Wanderungsrichtung, kann auch von männlichen (wandern zu ihren Partnern, die stationär bleiben) und weiblichen Kernen gesprochen werden], bilden funktionell eine Einheit (n+n), ohne miteinander zum diploiden Kern (2n) verschmolzen (K!) zu sein; sie teilen sich mitotisch synchron, um damit zwei Dikaryen zu bilden, sich zu verdoppeln. Im Entwicklungskreislauf erfolgt zwar zunächst die Plasmogamie (P!), doch die Karyogamie (K!) zum diploiden (2n) Kern wird erst kurz vor der Meiose (R!) vollzogen. Man spricht hier auch von verzögerter Karyogamie. Dikaryen treten bei Taphrinomycotina, Pezizomycotina, Pucciniomycotina, Ustilaginomycotina und Agaricomycotina auf.
[26] Diploid: Zellkerne mit doppeltem Satz zusammenpassender, homologer, Chromosomen; ausgedrückt mit 2n
[27] Meiose, meiotisch, R!: Meiose dient der Reduktion eines diploiden Chromosomensatzes zu haploiden Sätzen. Dabei werden einander entsprechende Chromosomen, im Kern sich dann mittig in einer Ebene gegenüberstehend, gepaart und anschließend in entgegengesetzter Richtung („polwärts“) separiert. Dieser Vorgang wird auch als Reduktionsteilung (oft abgekürzt als R! und zugleich stellvertretend für die ganze Meiose verwendet) bezeichnet. Da die voneinander getrennten haploiden Chromosomen schon zu Chromatiden verdoppelt sind, schließt sich an die Reduktionsteilung noch eine mitotische Teilung an, so dass vier haploide Kerne letztlich vorliegen.
[28] Zygote: Diploide Zelle, die nach der Verschmelzung zweier haploider Kerne, im Zuge der sexuellen Fortpflanzung entstand
[29] Haploid: Zellkerne mit einfachem Chromosomensatz; ausgedrückt als n
[30] Sporangium: Behälter, in dem Sporen asexuell oder sexuell gebildet werden
[31] Gameten: Für sexuelle Fortpflanzung vorgesehene haploide Zellen
[32] Gametangien: Behälter für Gameten; oder für Zellkerne, die, ohne Gameten zu bilden, dazu bestimmt sind, früher oder später über Karyogamie der sexuellen Fortpflanzung zu dienen
[33] Algen: Eine organismenreichübergreifende Bezeichnung für überwiegend im Wasser lebende, grüne Thallophyten
[34] Dikaryotische Phase: Ein Abschnitt im Lebenskreislauf, der durch Dikaryen ausgezeichnet ist
[35] Pezizomycotina und Basidiomycota
[36] Halicystis
[37] Xylan (Bryopsidales): Polymer aus β-1,3 glycosidisch verknüpften Xylose-Einheiten
[38] Cellulose: Unverzweigte Ketten aus Glucose in β-1,4-Verknüpfung; wobei der 6C-Zucker Glucose in Ring-Form geschrieben, das C1 der Aldehydgruppe ist [CH2OHCHOHCHOHCHOHCHOHCHO], davon aus gerechnet ist der vierte Kohlenstoff das C4 ist. In Ringform geschrieben weist die OH-Gruppe des C1 nach oben, wie auch die frei gebliebene CH2OH-Gruppe. Die OH-Gruppen wechseln von 1 bis 4 die Stellung: C1 nach oben, C2 nach unten, C3 noch oben, C4 nach unten, an C5 hängt die nach oben stehende CH2OH-Gruppe.
[39] Polysaccharide: Vielfachzucker aus einer großen, unbestimmbaren Zahl von Einfachzuckern (Monosacchariden)
[40] Derbesia
[41] Mannan: Polymere aus β-1,4 glycosidisch verknüpfen Mannose-Einfachzucker
[42] Bryopsis plumosa: Flaumige Moosalge (Bryopsidaceae – Bryopsidales – Ulvophyceae – Chlorophyta – Plantae –…)
[43] Codium fragile: Toten-Manns-Finger, Meergeweih (Bryopsidaceae – Bryopsidales – Ulvophyceae – Chlorophyta – Plantae – …)
[44] Thallus: Körper von Pflanzen (Pilzen), die nicht in Spross, Blatt und Wurzel gegliedert sind
[45] Gametischer Kernphasenwechsel: Meiose erfolgt im Zuge der Gametenbildung; nur Gameten sind haploid alle anderen Zellen sind diploid
Eingestellt am 21. Februar 2026
.

Halicystis (Gametophyt) von Derbesia (Sporopohyt)
Derbesia marina, in Kathy Ann Miller (ed.), 2025 California Seaweeds eFlora, http://ucjeps.berkeley.edu/seaweedflora/eflora_display.php?tid=47 [accessed on October 12, 2025]
Autor: Kathy Ann Miller (ed.), 2025 California Seaweeds eFlora
Lizenz: angefragt, 13.10.25, e-mail, Dr. Gilles, Director of The University and Jepson Herbaria, Berkeley University
Derbesia marina
{kind=link}
Autor: Philippe Bourjon
Lizenz: Creative Commons Attribution-Share Alike 4.0 International license; unverändert
Eingestellt am 21. Februar 2026
.
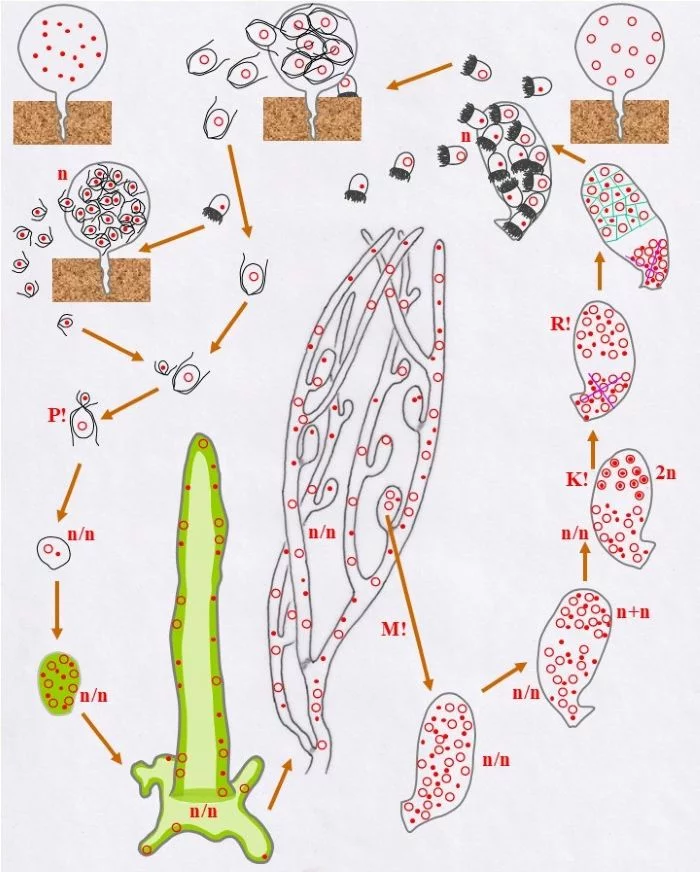
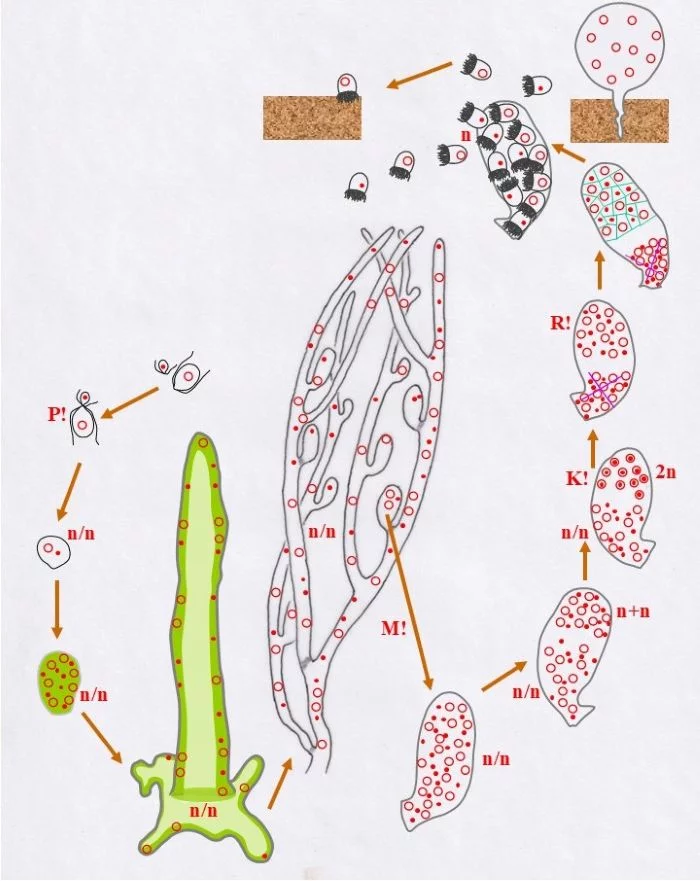
Entwicklungszyklus von Derbesia marina (Derbesia – Halicystis); (Tusche- und ppt-generiert; Reinhard Agerer)
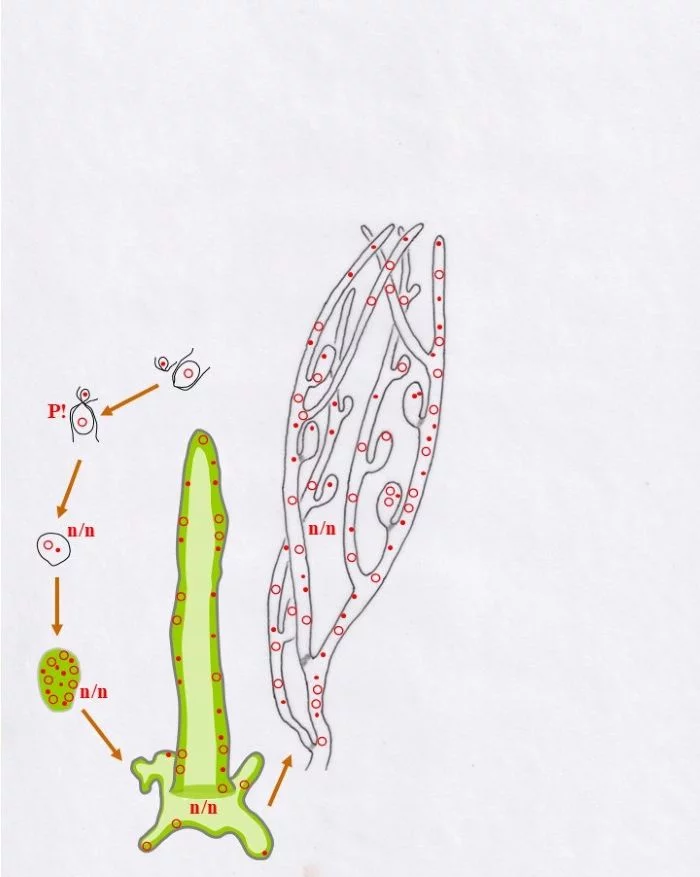
Zwei unterschiedlich große Gameten, ein kleiner männlicher (Punkt-Kern) und ein größerer weiblicher (Kringel-Kern) verschmelzen miteinander Kopf an Kopf (P!), verschmelzen die beiden haploiden Kerne aber nicht; die Karyogamie findet noch nicht statt, die Kerne bleiben als selbständige Einheiten erhalten. – Ob dies als Beginn der sporophytischen Phase zu betrachten ist oder nicht, bleibt dahingestellt, dazu mehr im Anschluss und am Ende der Legende. –
Die so vorliegende Zelle vermehrt mitotisch ihre Kerne; wobei keine Koppelung der beiden konträrgeschlechtlichen Kerne zu erkennen ist, sie bleiben offenbar als selbständige Einheiten erhalten. Damit entsteht eine junge heterokaryotische Pflanze, was hier mit n/n ausgedrückt wird. Diese vergrößerte vielkernige Zelle wächst zu einem ersten siphonalen Schlauch aus, der sich am Substrat mit einem plattenförmigen, unregelmäßigen Fuß verankert. Die Kerne liegen randlich im Protoplasten der zentral vakuolisierten Zelle (helleres Grün) in unregelmäßiger Anordnung, also ohne, dass die konträrgeschlechtlichen Kerne (Punkt-Kerne, bzw. Kringel-Kerne), so der Anschein, miteinander in engerer körperlicher oder funktioneller Beziehung stehen. Dieser erste Schlauch wächst zu einem verzweigten System, das ebenso heterokaryotisch ist. Daran entstehen ovale Zellen, die, nachdem sie ihre vorgegebene Größe erreichten, mit einem Pfropf abgetrennt werden; keine Querwände! (hellgraue Querbereiche). Solche Pfropfen werden auch ab und an in das Schlauchsystem eingezogen.
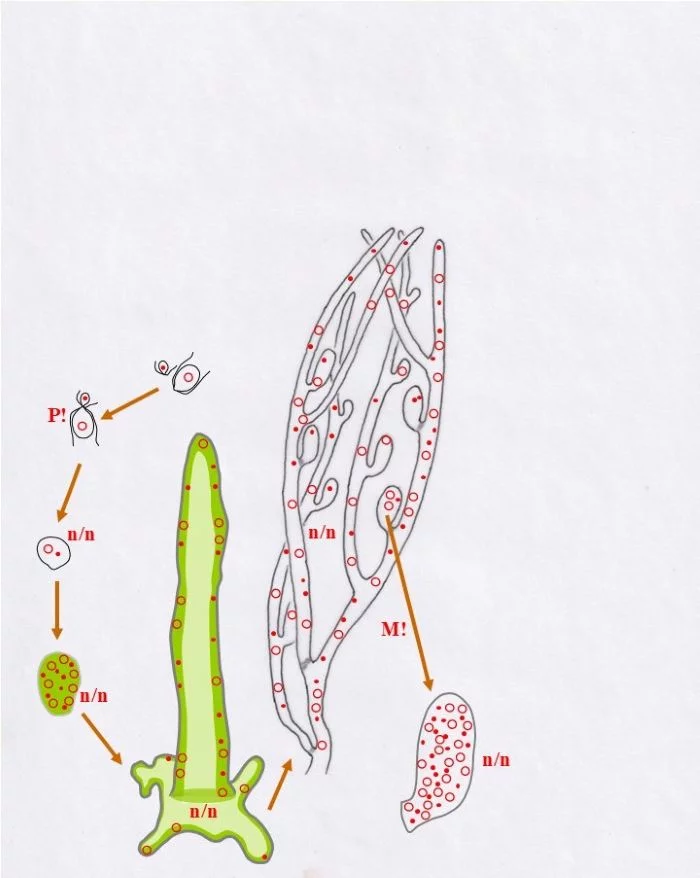
Diese Behälter („Vesikel“) sind durch mehrfache Mitosen (M!) vielkernig und heterokaryotisch (n/n)
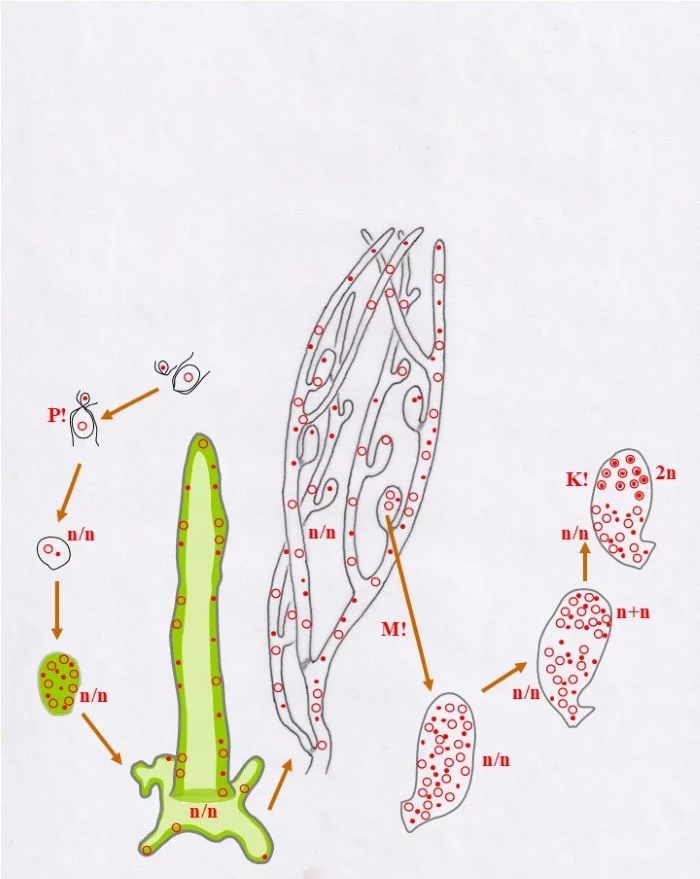
Sie durchlaufen nun, was die Kerne betrifft, eine spezielle interne Differenzierung: Im oberen Bereich paaren sich die Kerne zu Dikaryen (n+n), wobei sich konträrgeschlechtliche Kerne einander zuordnen (Punkt-Kerne, bzw. Kringel-Kerne), sich damit koppeln, was einem typischen Dikaryon entspricht. Die Dikaryen vollziehen die Karyogamie (K!), wodurch diploide (2n) Zellkerne entstehen. Die übrigen, die ungepaarten, Kerne werden funktionslos, bleiben im heterokaryotischen (n/n) unteren Bereich.
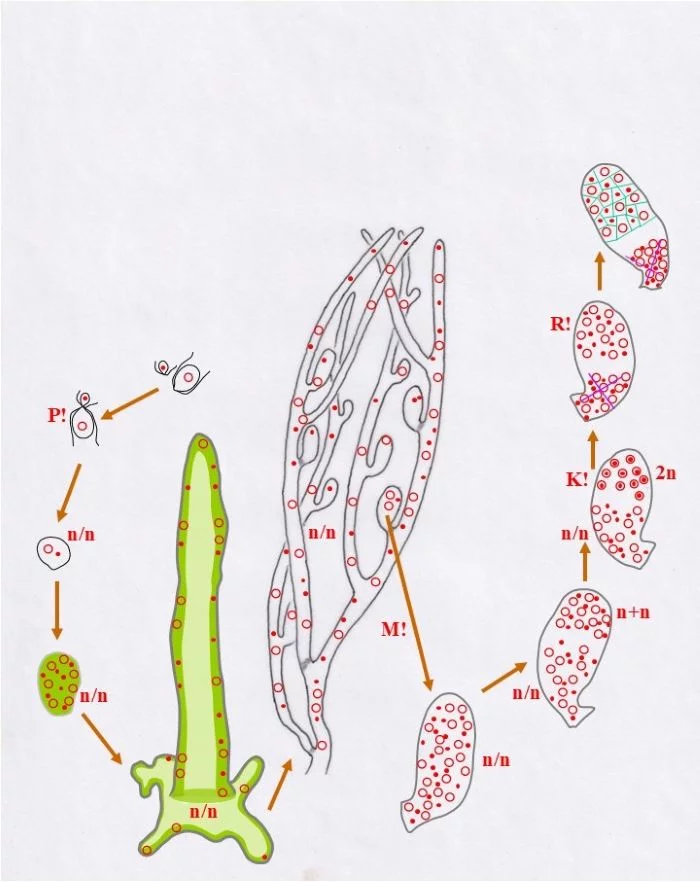
Die diploiden Zellkerne vollziehen nun die Meiose (R!) und grenzen sich voneinander ab (grüne Linien); so entstehen einkernige haploide Zellen unterschiedlichen Geschlechts männliche (Punkt-Kern-Zellen und weibliche Kringel-Kern-Zellen); die heterokaryotische Kernansammlung bleibt davon unberührt.
Diese Zellen bilden haploide Stephanosporen beiderlei Geschlechts (Punkt-Kerne, bzw. Kringel-Kerne) in einem nun als Zoosporangium erkennbaren Behälter. Diese Stephanosporen setzen sich auf dem Substrat fest und wachsen zu vielkernigen haploiden Blasen (Halicystis) aus.
Wie dies mit weiblichen Stephanosporen geschieht, so auch mit männlichen. Beide Behälter sind Gametangien (weibliche und männliche leben somit getrennt voneinander).
Sie entlassen ungleichgroße Gameten, große weibliche und kleine männliche, durch einen Porus der Gametangienwand.
Große weibliche (Kringel-Kern) und kleine männliche Gameten (Punkt-Kern) finden einander und vollziehen Plasmogamie (P!). Damit ist der Entwicklungskreislauf geschlossen.
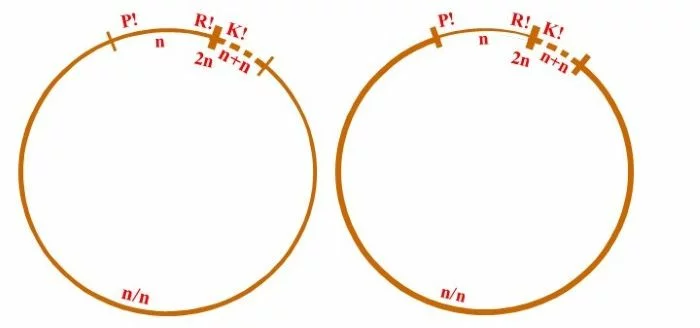
Allgemein wird der Entwicklungskreislauf von Derbesia – Halicystis als heterophasisch [zwei verschiedene Genrationen: haploider Gametophyt (blasig, Halicystis) und diploider Sporophyt (siphonal-fädig, Derbesia)], und heteromorph (Gametophyt und Sporophyt unterschiedlicher Gestalt) bezeichnet; wobei Gametophyten als Männlich und Weiblich getrennt auftreten. Unterschiedlich lassen sich aber die Anteile von Gametophyt und Sporophyt interpretieren: Rechts: In diesem Schema beginnt mit der Plasmogamie (P!) die sporophytische heterokaryotische (n/n) Phase, die sich mit der Dikaryophase (n+n), somatische und funktionelle Kopplung zweier konträrgeschlechtlicher Kerne, seinem Ende entgegengeht und mit der Karyogamie (K!) und der resultierenden diploiden Zygote (2n) abschließt. Meiose (R!) läutet die gametophytische Phase ein, die mit der Verschmelzung der Gameten (P!) endet. In dieser Darstellung zählt die heterokaryotische Phase (n/n) zum Sporophyten, wobei zu bemerken ist, die Kerne sind zwar kompatibel, doch haben sie miteinander offenbar nichts zu tun. Das Zusammenbringen der nicht verschmelzenden, sonst aber unabhängigen Kerne wird hier als Kriterium für die Entstehung des Sporophyten genommen, wie dies von Phykologen gesehen wird. Links: Hier ist die mögliche Sichtweise eines Mykologen dargestellt, der sich an Lebenszyklen der Dikarya orientiert, wie sie bei Pezizomycetes und Basidiomycota verwirklicht sind. Dort entsteht der Sporophyt mit dem Zusammenbringen und der Koppelung (Dikaryotisierung, n+n) konträrgeschlechtlicher Kerne und die sporophytische Phase endet mit der Karyogamie. Dort markieren nach Karyogamie und Meiose mit trichalen Fäden keimende Sporen die haploide, homokaryotische (n), trichale, gametophytische Phase. Doch können auch diese Fäden mehrkernig, homokaryotisch (n) oder heterokaryotisch (n/n) sein, ohne sexuell kompatible Kerne zu enthalten, was jedoch nicht auszuschließen ist. Das Zusammenbringen nicht verschmelzender, sonst aber gekoppelter, sexuell verträglicher Kerne wird dort als Kriterium für die Entstehung des Sporophyten genommen, also der Beginn der Diakryophase (n+n).
Fazit: Je nach Betrachtungsweise zeigt Derbesia eine kurze haploide gametophytische Phase: n zwischen R! und P! (rechter Kreislauf) oder eine lange, werden n und n/n (zwischen R! und Dikaryotisierung = Kernkopplung) zusammen als haploide Phase genommen. Nur die dikaryotische Phase (n+n) gälte im zweiten Fall (linker Kreislauf) als sporophytisch. – Alles Ansichtssache!
Nach van den Hoek et al. (1995/2002), Fig. 24.8, S. 429.
In den nachfolgenden zehn Abbildungen ist der Entwicklungszyklus einer Powerpoint-Präsentation ähnlich aufgebaut.
Eingestellt am 21. Februar 2026
.
Zwei unterschiedlich große Gameten, ein kleiner männlicher (Punkt-Kern) und ein größerer weiblicher (Kringel-Kern) verschmelzen miteinander Kopf an Kopf (P!), verschmelzen die beiden haploiden Kerne aber nicht; die Karyogamie findet noch nicht statt, die Kerne bleiben als selbständige Einheiten erhalten. – Ob dies als Beginn der sporophytischen Phase zu betrachten ist oder nicht, bleibt dahingestellt, dazu mehr im Anschluss und am Ende der Legende. –
Die so vorliegende Zelle vermehrt mitotisch ihre Kerne; wobei keine Koppelung der beiden konträrgeschlechtlichen Kerne zu erkennen ist, sie bleiben offenbar als selbständige Einheiten erhalten. Damit entsteht eine junge heterokaryotische Pflanze, was hier mit n/n ausgedrückt wird. Diese vergrößerte vielkernige Zelle wächst zu einem ersten siphonalen Schlauch aus, der sich am Substrat mit einem plattenförmigen, unregelmäßigen Fuß verankert. Die Kerne liegen randlich im Protoplasten der zentral vakuolisierten Zelle (helleres Grün) in unregelmäßiger Anordnung, also ohne, dass die konträrgeschlechtlichen Kerne (Punkt-Kerne, bzw. Kringel-Kerne), so der Anschein, miteinander in engerer körperlicher oder funktioneller Beziehung stehen. Dieser erste Schlauch wächst zu einem verzweigten System, das ebenso heterokaryotisch ist. Daran entstehen ovale Zellen, die, nachdem sie ihre vorgegebene Größe erreichten, mit einem Pfropf abgetrennt werden; keine Querwände! (hellgraue Querbereiche). Solche Pfropfen werden auch ab und an in das Schlauchsystem eingezogen.
Diese Behälter („Vesikel“) sind durch mehrfache Mitosen (M!) vielkernig und heterokaryotisch (n/n)
Sie durchlaufen nun, was die Kerne betrifft, eine spezielle interne Differenzierung: Im oberen Bereich paaren sich die Kerne zu Dikaryen (n+n), wobei sich konträrgeschlechtliche Kerne einander zuordnen (Punkt-Kerne, bzw. Kringel-Kerne), sich damit koppeln, was einem typischen Dikaryon entspricht. Die Dikaryen vollziehen die Karyogamie (K!), wodurch diploide (2n) Zellkerne entstehen. Die übrigen, die ungepaarten, Kerne werden funktionslos, bleiben im heterokaryotischen (n/n) unteren Bereich.
Die diploiden Zellkerne vollziehen nun die Meiose (R!) und grenzen sich voneinander ab (grüne Linien); so entstehen einkernige haploide Zellen unterschiedlichen Geschlechts männliche (Punkt-Kern-Zellen und weibliche Kringel-Kern-Zellen); die heterokaryotische Kernansammlung bleibt davon unberührt.
Diese Zellen bilden haploide Stephanosporen beiderlei Geschlechts (Punkt-Kerne, bzw. Kringel-Kerne) in einem nun als Zoosporangium erkennbaren Behälter. Diese Stephanosporen setzen sich auf dem Substrat fest und wachsen zu vielkernigen haploiden Blasen (Halicystis) aus.
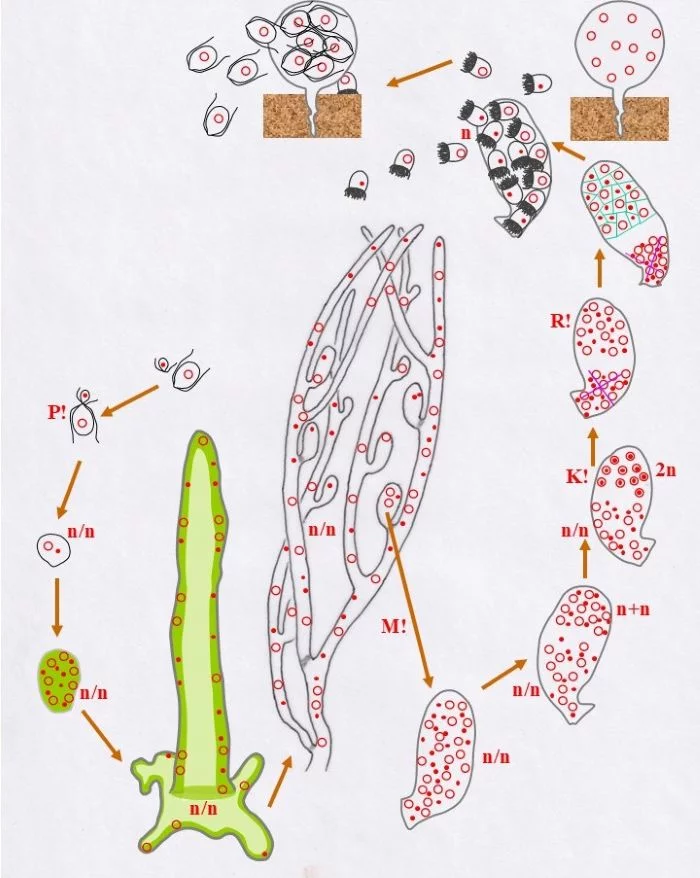
Wie dies mit weiblichen Stephanosporen geschieht, so auch mit männlichen. Beide Behälter sind Gametangien (weibliche und männliche leben somit getrennt voneinander).
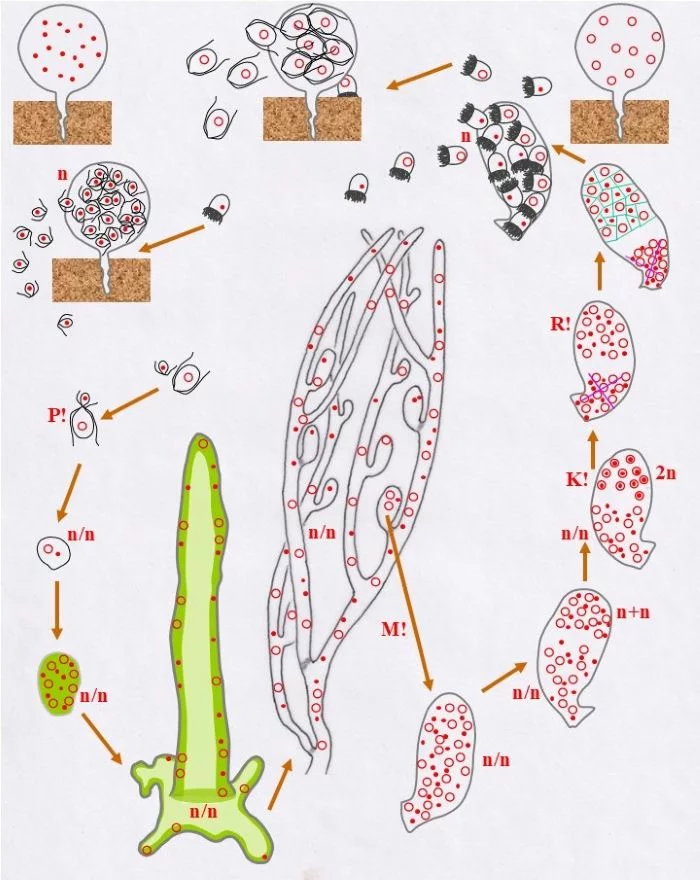
Sie entlassen ungleichgroße Gameten, große weibliche und kleine männliche, durch einen Porus der Gametangienwand.
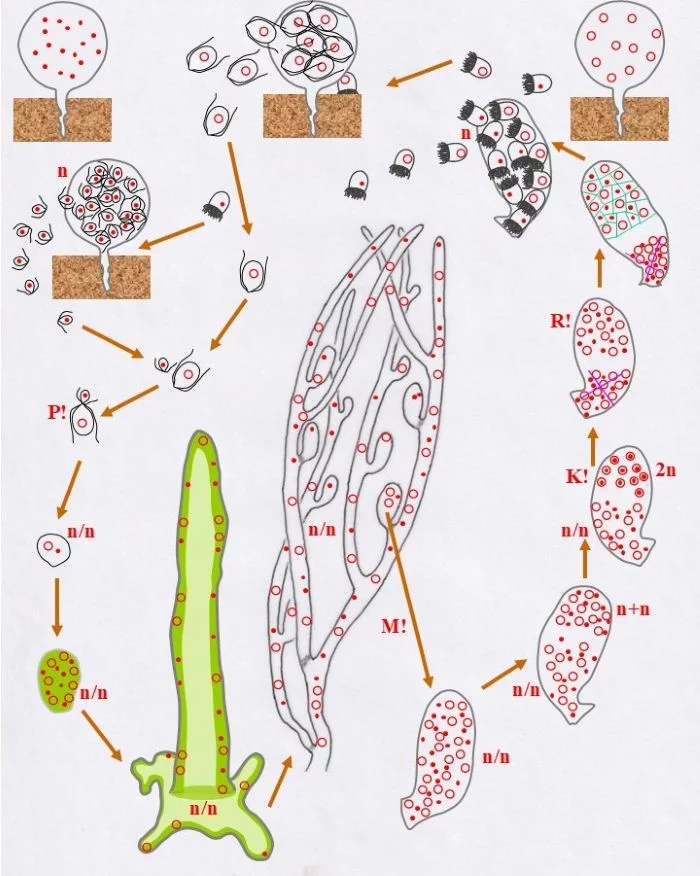
Große weibliche (Kringel-Kern) und kleine männliche Gameten (Punkt-Kern) finden einander und vollziehen Plasmogamie (P!). Damit ist der Entwicklungskreislauf geschlossen.
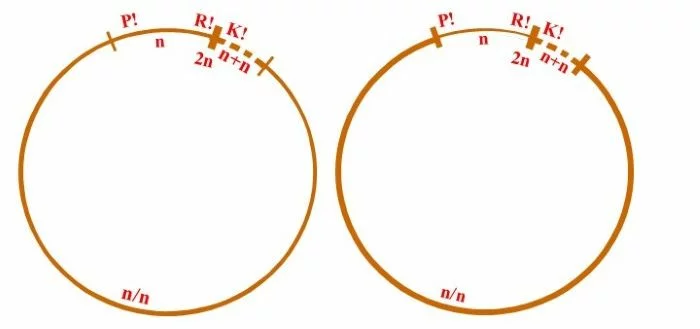
Allgemein wird der Entwicklungskreislauf von Derbesia – Halicystis als heterophasisch [zwei verschiedene Genrationen: haploider Gametophyt (blasig, Halicystis) und diploider Sporophyt (siphonal-fädig, Derbesia)], und heteromorph (Gametophyt und Sporophyt unterschiedlicher Gestalt) bezeichnet; wobei Gametophyten als Männlich und Weiblich getrennt auftreten. Unterschiedlich lassen sich aber die Anteile von Gametophyt und Sporophyt interpretieren: Rechts: In diesem Schema beginnt mit der Plasmogamie (P!) die sporophytische heterokaryotische (n/n) Phase, die sich mit der Dikaryophase (n+n), somatische und funktionelle Kopplung zweier konträrgeschlechtlicher Kerne, seinem Ende entgegengeht und mit der Karyogamie (K!) und der resultierenden diploiden Zygote (2n) abschließt. Meiose (R!) läutet die gametophytische Phase ein, die mit der Verschmelzung der Gameten (P!) endet. In dieser Darstellung zählt die heterokaryotische Phase (n/n) zum Sporophyten, wobei zu bemerken ist, die Kerne sind zwar kompatibel, doch haben sie miteinander offenbar nichts zu tun. Das Zusammenbringen der nicht verschmelzenden, sonst aber unabhängigen Kerne wird hier als Kriterium für die Entstehung des Sporophyten genommen, wie dies von Phykologen gesehen wird. Links: Hier ist die mögliche Sichtweise eines Mykologen dargestellt, der sich an Lebenszyklen der Dikarya orientiert, wie sie bei Pezizomycetes und Basidiomycota verwirklicht sind. Dort entsteht der Sporophyt mit dem Zusammenbringen und der Koppelung (Dikaryotisierung, n+n) konträrgeschlechtlicher Kerne und die sporophytische Phase endet mit der Karyogamie. Dort markieren nach Karyogamie und Meiose mit trichalen Fäden keimende Sporen die haploide, homokaryotische (n), trichale, gametophytische Phase. Doch können auch diese Fäden mehrkernig, homokaryotisch (n) oder heterokaryotisch (n/n) sein, ohne sexuell kompatible Kerne zu enthalten, was jedoch nicht auszuschließen ist. Das Zusammenbringen nicht verschmelzender, sonst aber gekoppelter, sexuell verträglicher Kerne wird dort als Kriterium für die Entstehung des Sporophyten genommen, also der Beginn der Diakryophase (n+n).
Fazit: Je nach Betrachtungsweise zeigt Derbesia eine kurze haploide gametophytische Phase: n zwischen R! und P! (rechter Kreislauf) oder eine lange, werden n und n/n (zwischen R! und Dikaryotisierung = Kernkopplung) zusammen als haploide Phase genommen. Nur die dikaryotische Phase (n+n) gälte im zweiten Fall (linker Kreislauf) als sporophytisch. – Alles Ansichtssache!
Nach van den Hoek et al. (1995/2002), Fig. 24.8, S. 429.
Eingestellt am 21. Februar 2026
.
Endpunkt erreicht